gordon0030@yandex.ru

Живая и неживая материя
 21.05.2003
21.05.2003  50:06
50:06
 Стенограмма эфира
Стенограмма эфираСуществует ли общая закономерность в эволюции живой и неживой материи? Можно ли подходить к оценке живого с позиции физики и рассматривать формирование жизни как высшее проявление происходящих в природе процессов самоорганизации? О том, как формировалась живая материя во Вселенной, — биолог Мстислав Крылов и физик Михаил Либенсон.
Участники:
Мстислав Владимирович Крылов — доктор биологических наук, научный сотрудник Зоологического Института РАН
Михаил Наумович Либенсон — доктор
Обзор темы
Известно, что в древнейших
Вселенная, которой свойственны неравновесность и, частично, необратимость, постоянно расширяется, а уровень ее организации усложняется. В неравновесных системах универсальный способ передачи энергии от одной химической реакции к другой осуществляется путем сопряжения этих реакций через общий промежуточный продукт. «Разрешающее» условие для самоорганизации — существование положительных обратных связей в системе и «каталитических» эффектов. Те же химические реакции могут ускоряться или замедляться одним из ее продуктов. Стационарному состоянию неравновесной системы (в условиях, препятствующих достижению равновесного состояния) соответствует стремление к минимальному производству энтропии.
Если к оценке живого подходить с позиции физики неравновесных процессов, то жизнь укладывается в рамки естественного порядка и может быть определена как высшее проявление происходящих в природе процессов самоорганизации. Живая клетка — открытая неравновесная система, находящаяся в стационарном состоянии, при котором скорость притока вещества и энергии соответствует скорости оттока. Для живой материи характерно усложнение организации: синтез больших молекул, молекулярных комплексов, клеток и многоклеточных организмов — это мощные антиэнтропийные факторы. Следовательно, усложнение живых организмов в процессе эволюции определяется законами термодинамики неравновесных процессов.
Функционирование экосистем определяется кооперативными взаимоотношениями между организмами. Отклонения от требований, предъявляемых системой, выбраковываются естественным отбором и борьбой за существование. Изменение условий инициирует самоорганизацию генома.
Таким образом, в эволюции живой и неживой материи прослеживается общая закономерность: сложным формам предшествуют менее сложные. Химические элементы образовались после появления протонов, нейтронов и электронов, сложные молекулы не могли сформироваться раньше химических элементов, эвкариотам предшествовали прокариоты, многоклеточным формам жизни — одноклеточные, амфибиям — рыбы, рептилиям — амфибии и т. д. Следовательно, «разрешающими» условиями для появления более сложных форм материи, служит предшествующее им образование менее сложных. «Старое» способствует возникновению «нового», «новое» изменяет условия, в которых существование «старого» становится затрудненным, и оно либо гибнет, либо уходит с авансцены, либо преобразуется. Эволюция осуществляется дискретно по эстафетному принципу. При этом сходные или одинаковые формы материи, в том числе и живой, возникают везде, где есть «разрешающие» условия, которые создаются самой материей и являются ее частью. Путь создания живого — единственный во Вселенной, как и путь построения протонов, нейтронов, атомов водорода, гелия и остальных атомов.
И наконец, последний вывод из существования этой иерархии: формирование жизни — естественный этап развития материи, поэтому Землю нельзя считать единственным обитаемым космическим телом.
Современная картина мира, в основу которой положены необратимые процессы, существование обратных связей и нелинейность, дает возможность выработать логически целостный подход к пониманию механизмов эволюции живой и неживой материи. Живые организмы и неживая природа подчиняются одним и тем же концептуальным законам физики и химии.
Рассмотрим сначала в общих чертах эволюцию ранней Вселенной. Под этим обычно понимается эпоха, соответствующая возрасту от 10−43 секунды до 3 минут от начала истории. Согласно общепризнанной теории Большого Взрыва, рождение классического пространства — времени произошло из сингулярности, причем в момент рождения Вселенной плотность и температура вещества достигали планковских значений 1093 г/см³
Таким образом, расширяющейся Вселенной свойственна неравновесность и связанные с ней усложнение и возможная необратимость. Это способствует неравновесности и необратимости многочисленных процессов в природе, включая жизнь. Неравновесность и необратимость могут быть источниками порядка, согласованности и повышения уровня организации.
К настоящему времени накопилось много данных о самоорганизующихся неравновесных системах различных уровней сложности, начиная от самоорганизации, происходящей в неорганических химических системах, где участвующие молекулы просты до морфогенеза сложных живых систем. Например, возникновение химических часов — химических реакций с согласованным периодическим изменением концентрации реагентов. В случае химических часов все молекулы изменяют свое химическое тождество одновременно, через правильные промежутки времени. Такого типа связь широко распространена в живых системах.
При «разрешающих» условиях олигомерные белки, например, гемоглобин и аллостерические ферменты обнаруживают способность к самосборке. При этом выявляется высокая специфичность взаимодействующих субъединиц. Гемоглобин и альдолаза в смеси сохраняют молекулярную идентичность, даже смесь натцвных гемоглобинов, выделенных из двух видов млекопитающих, не образует гибридных молекул. Информация о структурной организации полипептидов содержится в аминокислотной последовательности и может сохраняться даже при денатурации. Изменение условий, например, подкисление раствора альдолазы приводит к ее диссоциации на неактивные субъединицы. Если затем постепенно довести рН раствора до 7,0, то субъединицы самопроизвольно восстанавливают свою нативную конфирмацию.
Самоорганизация распространяется и на системы более высокого порядка, так формируются мультиферментные комплексы, происходит самосборка мембранных структур, рибосом, вируса табачной мозаики. При изучении морфогенеза бактериофага Т4 удалось наблюдать процессы формирования сложной трехмерной структуры на основе «одномерной» информации, содержащейся в аминокислотных последовательностях белковых субъединиц. Самоорганизация морфогенеза более сложных структур контролируется специальными морфогенами.
В состояниях, далеких от равновесия, происходят и другие спонтанные перераспределения материи. В неравновесных системах универсальный способ передачи энергии от одной химической реакции к другой осуществляется путем сопряжения этих реакций через общий промежуточный продукт; именно таким образом происходит перенос энергии почти во всех обменных реакциях в живых клетках. «Разрешающим» условием для прохождения процесса самоорганизации является существование каталитических эффектов, а также существование с самого начала или возникновение в процессе реакции системы обратных связей — положительных и отрицательных. Ускорение или замедление химических реакций может осуществляться одним из ее продуктов. Такие явления называют автокаталитическими и автоингибирующими процессами. В неорганической химик эти процессы встречаются редко, но как показали исследования молекулярной биологии, они составляют основу жизни. Именно такие процессы позволяют понять, каким образом совершается переход от небольших цепочек нуклеиновых кислот к сложным организмам.
Можно привести множество примеров самоорганизации в окружающем нас мире. Это и образование причудливых по форме и структуре облаков, и специфические структуры вихрей, торнадо, циклонов, а также узоров, которые рисует мороз на стекле и т. п. Процессы самоорганизации сопровождают формирование новых фаз вещества при плавлении и кристаллизации. Очень характерны в этом отношении и специфические структуры, возникающие на поверхности самых различных материалов под воздействием интенсивного лазерного излучения.
Живая клетка — открытая неравновесная система, находящаяся в стационарном состоянии, при котором скорость «притока» вещества и энергии соответствует скорости «оттока». Стационарному состоянию неравновесной системы (в условиях, препятствующих достижению равновесного состояния) соответствует стремление к минимальному производству энтропии. Для живой материи характерно усложнение организации: синтез больших молекул, молекулярных комплексов, клеток и многоклеточных организмов — это мощные антиэнтропийные факторы. Таким образом, живой организм — это наиболее упорядоченное состояние открытой неравновесной, находящейся в стационарном состоянии системы, для которой характерна минимальная скорость возрастания энтропии. Следовательно, усложнение живых организмов в процессе эволюции определяется законами термодинамики неравновесных процессов и самоорганизации. Если к оценке живого подходить с позиции физики неравновесных процессов, то жизнь укладывается в рамки естественного подхода и может быть определена как высшее проявление, происходящих в природе процессов самоорганизации. Ископаемые формы жизни, обнаруженные на ранней Земле (800 млн лет назад), могут служить серьезным аргументом в пользу идей о спонтанной самоорганизации живого в «разрешающих» условиях.
Различные количественные сочетания протонов, нейтронов и электронов сопровождаются качественным изменениям и дают начало различным атомам. Последующая форма организации материи — химический элемент, это совокупность атомов с одинаковым зарядом атомных ядер и одинаковым числом электронов в ядерной оболочке.
В эволюционном ряду элементарные частицы — атомы, химические элементы — молекулы, очень сильно возрастает число форм и возможных состояний материи. Формирование молекул из атомов, также как и формирование атомов из элементарных частиц происходит под влиянием сил взаимодействия и осуществляется только при определенных условиях, «разрешающих» пройти процессу, детерминированному природой этих форм материи. Так, например, условия, при которых возможен синтез ядер гелия из водорода —
Хорошо известно, что в зависимости от давления и температуры одни и те же химические элементы могут находиться в различных состояниях. Так, азот при обычных условиях — газ, при −147 °C — жидкость; кислород при обычных условиях — газ, при −182,9 °С — жидкость, при −218,7 °С образует кристаллы.
Таким образом, появление новых форм и состояний материи определяется изначально присущими материи определенными свойствами, детерминирующими ее последующие превращения, наличием взаимодействий между различными формами материи, появлением новых качеств у вновь сформировавшихся форм материи и условиями, «разрешающими» прохождению процессов эволюции. Начальные условия и процессы эволюции не могут быть независимыми. Именно начальные условия определили наличие на Земле химических элементов (около 22), «пригодных» для построения биомолекул, массу, плотность и орбиту вращения планеты вокруг Солнца. Эти особенности, «разрешившие» формирование жизни, являются важнейшим отличием Земли от других планет Солнечной системы, начальные условия на которых неблагоприятны для появления жизни.
Можно выстроить хронологию возникновения различных биохимических явлений на Земле. Примерно, она будет выглядеть так:
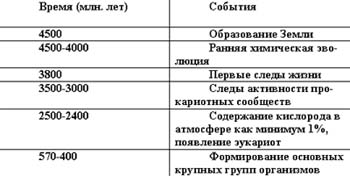
Эти данные позволяют предположить, что среди органических молекул, которые находились на ранней Земле, присутствовали молекулы, которые послужили строительными блоками для живых систем. Особенно важны для понимания эволюции живой материи на Земле находки в древнейших осадочных формациях
Начальным типом питания считают анаэробное брожение с субстратным фосфорилированием, использование неорганических доноров электрона, анаэробный фотосинтез (пурпурные и зеленые бактерии) и, наконец, аэробный фотосинтез с удалением кислорода, который резко меняет окислительную обстановку на Земле и создает необходимые условия для появления эукариот. Сложные эукариотные клетки, используя кислород, значительно увеличили производство энергии. Это было важным этапом в формировании многоклеточности. Таким образом, возникновению многоклеточности предшествовало появление кислорода в атмосфере Земли. Анаэробный организм не может быть слишком крупным
Накопление кислорода в атмосфере в результате фотосинтеза шло медленно, концентрация кислорода в атмосфере достигла 1% его содержания в современной атмосфере примерно
В сообществе разнородных организмов на первый план выходят кооперативные взаимоотношения, в которых определяющим существованием объекта оказывается его соответствие системе. Таким образом, экосистема является мощным двигателем развития, именно она предопределяет свойства объекта, входящего в нее. Г. Заварзин (2000) считает возможным поставить знак равенства между биогеохимической сукцессией и эволюцией биосферы.
Соответствие системе — непременное условие существования индивидуума. Отклонения от требований, предъявляемых системой, «выбраковываются» мощными помехоустойчивыми факторами — естественным отбором и борьбой за существование.
Анализ морфофункциональной организации генома указывает на способность его активно реагировать на изменения условий. К механизмам, на базе которых осуществляются эти процессы, можно отнести следующие:
• Эпигенетическая изменчивость — способность клетки целенаправленно переключаться с одной наследственной про граммы функционирования на другую, в зависимости от метаболической ситуации.
• Наличие в клетке сложной системы контроля над нарушениями структуры и функции ДНК, осуществляющейся на уровне репликации, транскрипции и трансляции. Все неточности и неисправности ДНК подвергаются репарации, если же ошибку не удается исправить, запускается система клеточной смерти. Этот механизм помехоустойчивости защищает геном от случайных, изменений и гарантирует биологическому объекту его соответствие экосистеме.
• Способность генетических элементов к мультипликации внутри генома, что ведет к качественному изменению последнего.
• Амплификационные перестройки генома связаны с умножением числа копий гена, при этом ген амплифицируется не один, а в составе сегментов хромосомы, иногда эти участки достигают миллионов оснований ДШС. Амплифицированные участки могут оставаться в исходной хромосоме или образовывать
• Мобильные генетические элементы (МГЭ), создающие новые конструкции, которые ведут к реорганизации генома и играют особую роль в эволюции, влияя на разнообразие организмов.
• Горизонтальный перенос генов — трансдукция, широко используется в генной инженерии, в естественных условиях ведет к преобразованию генома.
• Наличие одинаковых генов у далеко отстоящих в систематическом отношении организмов.
• Модульный принцип устройства генома, позволяющий быстро осуществлять построение новых конструкций, создающих огромные возможности для регуляции генных ансамблей.
Функционирующий геном представляет собой комплекс матричных систем, в которых поток информации идет в двух направлениях: от нуклеиновых кислот к белкам и от белков к нуклеиновым кислотам. Изменение условий инициирует самоорганизацию генома. Эти процессы невозможно отнести к случайным событиям. Ни один наблюдаемый процесс не может быть объяснен случайными событиями. Невозможно формулировать законы и делать обобщения о характере процесса или проверить его экспериментально, признавая возможность чистой случайности. Случайное событие и статистически случайное событие — два совершенно разных понятия. В природе не существует
Со временем уровень организации Вселенной неуклонно повышается. В эволюционных процессах живой и неживой материи прослеживается общая закономерность: сложным формам предшествуют менее сложные. Так, химическим элементам предшествует появление протонов, нейтронов и электронов, сложные молекулы не могут сформироваться раньше химических элементов, эукариотам предшествуют прокариоты, многоклеточным формам жизни — одноклеточные, амфибиям — рыбы, рептилиям — амфибии и т. д. Появление гелия становится возможным в различных областях Вселенной только после появления водорода, а эволюция млекопитающих могла начаться только после появления амфибий и рептилий. Формирование форм материи, служит предшествующее им образование менее сложных. «Старое» способствует возникновению «нового», «новое» изменяет условия, в которых существование «старого» становится затрудненным и оно либо гибнет, либо уходит с авансцены. Анаэробы способствовали появлению фотосинтезирующих
В первой половине двадцатого века была обнаружена независимая эволюция одинаковых тканей у членистоногих и позвоночных. Орган зрения — глаз возник независимо у разных групп организмов: книдарий, разных типов червей, моллюсков, членистоногих и позвоночных. Убедительные доказательства независимого формирования одинакового скелета можно проследить на примере строения раковин у простейших — фораминифер (Foraminifera) и головоногих моллюсков (Nautiloida), у двухстворчатых моллюсков (Bivalvia) и ракушечковых рачков (Ostracoda). Ряд не родственных между собой групп организмов независимо достиг артроподного уровня организации. Рептилийные признаки появились у разных групп амфибий асинхронно и независимо, формирование мамальных особенностей происходило различными темпами, независимо у различных групп териодонтов. Одинаковый набор типов листорасположения и листорассечения обнаружен у самых разных групп растений, от папоротников до высших покрытосемянных.
Путь создания живого — единственный во Вселенной, так же как и путь построения протонов, нейтронов, атомов водорода, гелия, углерода, кислорода. Формирование жизни — естественный этап развития материи, поэтому Землю нельзя считать единственным обитаемым космическим телом.
Библиография
Васильев А. Н. Эволюция Вселенной/Современное естествознание: Энциклопедия. Т. 4. Физика элементарных частиц. Астрофизика. М., 2000
Голубовский М. Д. Век генетики: эволюция идей и понятий. СПб., 2000
Гут А. Г., Стейнхард П. Д. Раздувающаяся Вселенная//В мире науки. 1984. № 7
Дуда В. И., Лебединский А. В., Кривенко В. В. Архибактерии в системе царств органического мира//Успехи микробиологии. 1985. Вып. 20
Еськов К. Ю. История Земли и жизни на ней. М., 2000
Жмур С. И., Розанов А. Ю., Горленко В. М. Следы древнейшей жизни в космических телах Солнечной системы//Природа. 1997. № 8
Заварзин Г. А. Фенотипическая систематика бактерий. М., 1974
Заварзин Г. А. Недарвиновская область эволюции//Вестник РАН. 2000. Т. 70. № 5
Кусакин О. Г., Дроздов А. Л. Филема органического мира. СПб., 1998. Ч. 2
Ленинджер А. Л. Биохимия. Молекулярные основы структуры и функции
Мейн С. В. О соотношении номогенетического и тихогенетического аспектов
Новиков И. Д. Инфляционная модель ранней Вселенной//Вестник РАН. 2001. Т. 71. № 10
Опарин А. И. Происхождение жизни. М., 1924
Опарин А. И. Возникновение жизни на Земле. М., 1957
Пригожин И., Стенгерс И. Порядок из хаоса. М., 1986; М., 2003
Сажин М. В. Космология ранней Вселенной/Современное естествознание: Энциклопедия. Т. 4. Физика элементарных частиц. Астрофизика. М., 2000
Спирин А. С. Биосинтез белков, мир РНК и происхождение жизни//Вестник РАН. 2001. Т. 71. № 4
Татаринов Л. П. Морфологическая эволюция териодонтов и общие вопросы филогенетики. М., 1976
Фолсом К. Е. Происхождение жизни: Маленький теплый водоем/Пер. с англ. Д. Б. Крипотина. М., 1982
Хокинг С. В. Краткая история времени от большого взрыва до черных дыр/Пер. с англ. Н. Смородинской. СПб., 2001
Холланд П.,
Эйген М. Самоорганизация материи и эволюция биологических молекул. М., 1973
Эйген М., Шустер П. Гиперцикл. Принципы самоорганизации молекул. М., 1982
Chetverina H. V., Demidenko A. A., Ugarov V.I., Chetverin A. B. Spontaneous rearrangements in RNA sequences//FEBS Letters. 1999. V. 450
Hall J. B. Evolution of the Prokaryotes//J. Theoret. Biol. 1971. V. 30
Krylov M. V., Libenson M. N. Continuum of evolutionary processes of living and nonliving matter//Proc. Zool. Inst. Russ. Acad. Sci. 2002. V. 296
Manton S. M. The Arthropoda habits functional morphology and evolution. Oxford, 1977
Zawarzin A. A. Der Parallelismus der Strukturen als ein Grundprinzip der Morphologie//Z. Wiss. Zoll. 1925. Bd 124.
Тема № 256
Эфир 21.05.2003
Хронометраж 50:06